 (Carregando Índice)... (Carregando Índice)... |
Você está em:
Inicial  acp-medicine Imunologia
acp-medicine Imunologia
Última revisão: 05/03/2012
Comentários de assinantes: 0
John R. David, MD
Richard Pearson Strong Professor, Department of Immunology and Infectious Diseases, Harvard School of Public Health, and Professor of Medicine, Harvard Medical School.
Cox Terhorst, PhD
Professor of Medicine, Harvard Medical School, and Chief, Division of Immunology, Beth Israel Deaconess Medical Center
Artigo original: David JR, Terhorst C. Organs and cells of the immune system. ACP Medicine. 2003;1-8.
[The original English language work has been published by DECKER INTELLECTUAL PROPERTIES INC. Hamilton, Ontario, Canada. Copyright © 2011 Decker Intellectual Properties Inc. All Rights Reserved.]
Tradução: Beatriz Rosário
Revisão técnica: Dr. Euclides Furtado de Albuquerque Cavalcanti
O sistema imunológico medeia a relação da pessoa com o ambiente microbiano. A imunidade envolve respostas inatas ou naturais e respostas altamente específicas adquiridas ou adaptáveis. A diferença essencial entre os dois tipos de imunidade está na forma pela qual os microrganismos são identificados. Na imunidade inata, os glicolipídeos e macromoléculas com padrões repetidos que são únicas nos organismos infecciosos são identificadas pelos receptores de superfície de células presentes nos macrófagos, células dendríticas, exterminadoras naturais (natural killers – NK) e células T NK (NKT), assim como pelo sistema complemento. Na imunidade adquirida, os linfócitos usam receptores de antígeno muito específicos para identificar agentes infecciosos e outros antígenos, de forma direta ou quando processados pelas células apresentadoras de antígenos (CAP), como as células dendríticas. Assim, existe um efeito recíproco entre a imunidade inata e a adquirida no nível da CAP. Uma vez que uma pessoa saudável tenha tido uma infecção causada por bactérias ou vírus, o sistema imunológico reconhece o patógeno e evita sua recorrência. Além disso, o sistema imunológico tem a incrível capacidade de distinguir antígenos, mesmo quando suas estruturas são muito semelhantes. Os linfócitos também podem reagir a autoantígenos, causando autoimunidade.
A resposta imunológica deve ser capaz de distinguir o que é próprio do organismo do que não é. Caso contrário, as células T e os anticorpos atacariam constantemente as células autólogas, componentes dos tecidos ou até bactérias comensais. Na década de 1950, Sir Frank Macfarlace Burnet propôs pela primeira vez que, no estágio pré-natal, a interação dos autoantígenos com linfócitos específicos para antígenos levaria à eliminação dos linfócitos autorreativos e, assim, à tolerância imunológica.1 Quando a tolerância imunológica não funciona, os anticorpos e as células sensibilizadas (reativas para antígenos) que são direcionadas contra autoantígenos provocam as doenças autoimunes [ver Tolerância imunológica e autoimunidade].
Existem dois grupos principais de linfócitos: as células T (também chamadas de linfócitos derivados do timo ou linfócitos T) e as células B (também chamadas de linfócitos derivados da medula óssea ou linfócitos B). As células T e B constituem entre 80 e 95% dos linfócitos do sangue periférico.
Estas células possuem um amplo poder de reconhecimento de antígeno. Duas características únicas sustentam esta capacidade:
uma família de células B de genes variáveis que, combinados, podem identificar um número quase infinito de antígenos;
uma família de células T de genes variáveis com apenas uma leve capacidade mais limitada. Nem as células T nem as células B constituem uma população homogênea de células; cada grupo inclui vários subgrupos que podem ser diferenciados pela região constante dos seus receptores, por conjuntos específicos de marcadores de superfícies expressos de maneira desenvolvida, sua localização nos órgãos linfoides e sua função. A ligação de combinações de anticorpos monoclonais aos receptores de superfície é atualmente a técnica mais específica usada para identificar os principais subconjuntos destas células.
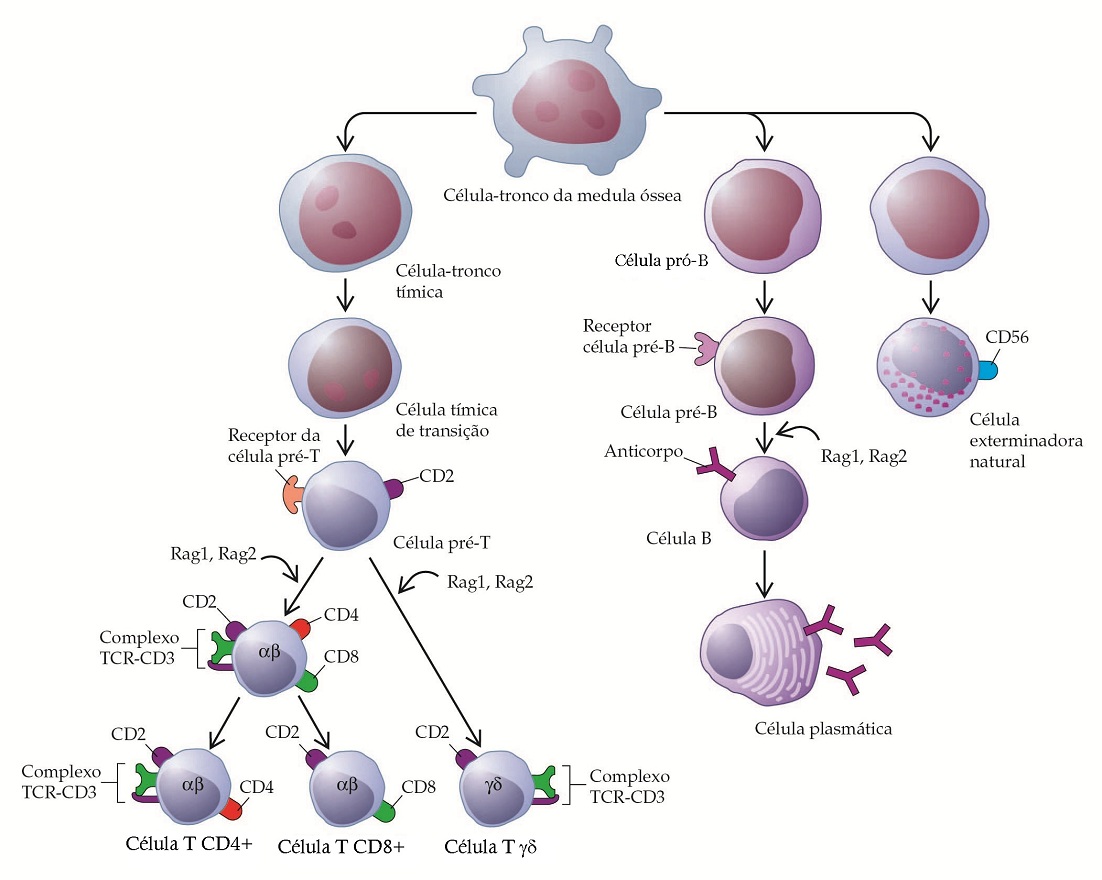
As células T maduras expressam os receptores de células T alfa-beta (TCR-alfa-beta) ou gama-delta (TCR-gama-delta) num complexo com as proteínas CD3. CD4 é expressa em 50 a 65% das células T alfa-beta periféricas e CD8 é expressa em 25 a 35% das células T alfabeta periféricas. Geralmente, células T gama-delta não apresentam CD4 nem CD8. Embora CD4 e CD8 sejam expressas juntas nos timócitos corticais, apenas uma ou outra é expressa nos subconjuntos complementares dos timócitos maduros e nas células T alfa-beta periféricas (células T CD4+ e CD8+) [ver Figura 1]. As células T CD4+ reconhecem o antígeno quando este é apresentado em associação com as moléculas da classe II (HLA-D e HLA-DR), o complexo maior de histocompatibilidade (CMH) ou em associação com CD1d, o último sendo células NKT. As células T CD8+ reconhecem o antígeno no contexto das moléculas classe I (HLA-A, HLA-B e HLA-C) do CMH [ver Mecanismos de resposta imunológica]. O contexto do reconhecimento do antígeno pela T gama-delta é desconhecido.
As células T CD4+ auxiliares (TH) podem ser também diferenciadas em células TH1 e TH2 com base nas citocinas que produzem.2 As células TH1 secretam interleucina-2 (IL-2) e interferon-gama (IFN-gama) que são importantes para a imunidade mediada por células. As células TH2 secretam IL-4, IL-5, IL-6, IL-10 e IL-13, que são críticas para a produção de anticorpos. As citocinas produzidas por cada um destes tipos de células também influenciam o outro tipo de células. Por exemplo, a IFN-gama produzida pelas células TH1 pode inibir a função das células TH2, enquanto a IL-10, que é secretada pelas células TH2, monócitos, macrófagos e células B, pode inibir a função das células TH1. Além de ter uma função auxiliadora, as células TH1 podem induzir cascatas inflamatórias levando a autoimunidade, como ocorre nas doenças inflamatórias no intestino e na artrite reumatoide.
A CD4 na superfície das células T auxiliares tem um papel importante na infecção pelo HIV. No estágio inicial da infecção, o vírus usa a CD4 como um correceptor, junto com CCR5, que por si só é um receptor para várias quimiocinas, incluindo RANTES (célula T normal expressa e secretada, regulada na ação), a proteína-1-alfa (MIP-1-alfa) inflamatória dos macrófagos e MIP-1-beta. Uma porcentagem dos indivíduos brancos é homozigota para um defeito no receptor CCR5 e é resistente a infecção pelo HIV. O receptor CCR4 da quimiocina, que é o receptor para o fator-1 estromal derivado de célula (SDF-1), está envolvido no estágio tardio da infecção pelo HIV.3,5

Figura 1. As linhagens de células do sistema imunológico e das células sanguíneas começam todas com a célula-tronco. As células-tronco que se diferenciaram para gerar células B residem na medula óssea, e as que produzem as células T migram da medula óssea para o timo. A maturação da célula T envolve a expressão progressiva dos marcadores de superfície de célula selecionados e a ativação de vários genes, incluindo os genes alfa, beta, gama e delta que codificam as cadeias que constituem os receptores alfabeta e gama-delta das células T (TCRs). As pessoas que não possuem a enzima Rag1 e Rag2 não progridem do estágio pré-célula T ou célula pré-B e, portanto, não possuem linfócitos. A seleção positiva e negativa de células T ocorre no estágio chamado duplo positivo (CD4+, CD8+). As células exterminadoras naturais se desenvolvem tanto na medula óssea quanto no timo.
As células B são precursoras das células produtoras de imunoglobulina (células plasmáticas) do sistema imunológico e são identificadas pela presença de imunoglobulina na sua superfície. Estas células com membrana de superfície positiva para imunoglobulina (SmIg+) constituem 5 a 15% dos linfócitos do sangue periférico. A maioria das células B possui IgM e IgD na sua superfície; cerca de um quarto de todas as células B possui apenas IgM ou IgD na sua superfície. Uma porcentagem de células B exibe IgG ou IgA.
Na superfícia das células B encontram-se o receptor 2 do complemento (CR2 ou CR21) que se liga a C3d/C3dg e ao vírus Epstein-Barr. Fc-gama-RIIb (CD32) é o principal receptor Fc nas células B, que está envolvido na ativação do linfócito B. Um marcador comum que é usado para identificar as células B é o CD19, que forma um grande complexo com CD21 e CD81 (alvo para o antígeno-1 antiproliferativo [TAPA-1]).6
As células B1, um subconjunto de células B, se desenvolvem precocemente e possuem uma vida muito longa. Os progenitores das células B são encontrados no fígado fetal e no omento embrionário, mas não na medula óssea adulta. As células B1 que expressam CD5 na sua superfície são chamadas de B1a, e as que não possuem CD5 são chamadas de B1b. As células B1 estão frequentemente associadas com a produção de autoanticorpos. Elas também produzem quantidade substancial de IL-10.7
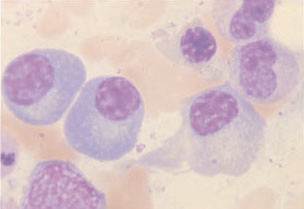
Sob a influência do antígeno, das células T e das células acessórias, as células B se diferenciam em células plasmáticas, as células maduras produtoras de anticorpos [Figura 1]. As células plasmáticas são maiores que os linfócitos e são caracterizadas por um núcleo circular excêntrico com heterocromática grosseira disposta num padrão de roda de carro. Estas células possuem um citoplasma altamente basofílico e um retículo endoplasmático bem desenvolvido, muitas vezes organizado em camadas paralelas concêntricas. Elas podem estar inchadas com material granular, que consiste em anticorpos produzidos por estas células [Figura 2]. Às vezes, uma ou mais cisternas endoplasmáticas estão distendidas por grandes inclusões chamadas de corpúsculos de Russel. Estes corpúsculos são agregados de moléculas de imunoglobulina incompletamente formadas. As células plasmáticas não apresentam mais a imunoglobulina de superfície. Elas também são células terminais, ou seja, elas não se dividem. É difícil distinguir os precursores imaturos das células plasmáticas, os plasmablastos dos linfoblastos e linfócitos grandes. Não é normal encontrar células plasmáticas no sangue periférico.
Figura 2. As células plasmáticas são células produtoras de anticorpos do sistema imunológico. Elas se diferenciam a partir das células B; possuem 6 a 20 µm de diâmetro, um núcleo excêntrico, um citoplasma altamente basofílico e uma área justanuclear proeminente e clara que contém o aparato de Golgi e o diplossomo.
As células exterminadoras naturais (natural killers – NK) são grandes linfócitos granulares que não possuem o complexo TCR-CD3 característico das células T ou o SmIg característico das células B. Uma célula-tronco derivada da medula óssea é a precursora das células T, B e NK [Figura 1]. In vitro, as células NK podem matar uma variedade de células tumorais e células infectadas com vírus de forma inespecífica, ou seja, eles não precisam da sensibilização prévia ou a presença de anticorpos para serem citotóxicos. Os grânulos contêm proteínas formadoras de poros que podem mediar a lise celular. As células NK expressam receptores inibitórios (KIR) que reconhecem as moléculas da classe I do CMH. Desta forma, as funções das células NK são inibidas por células que expressam a classe I do CMH, mas são ativadas por células que não possuem tal classe. As células NK humanas expressam um segundo grupo de receptores inibitórios, que incluem duas subunidades: uma subunidade NKG variável e a estrutura CD94 constante da superfície celular. Seu ligante é desconhecido. A IL-12 estimula as células NK a se proliferarem e produzirem IFN-gama, que é importante para várias reações imunológicas.8,9
Os monócitos pertencem ao sistema fagocítico mononuclear, antigamente chamado de sistema retículoendotelial. Eles são grandes células mononucleares que constituem 3% a 8% dos leucócitos do sangue periférico. Seu citoplasma é muito mais abundante do que o dos linfócitos. Geralmente, seu núcleo é excêntrico e tem formato oval ou de rim [Figura 3]. Os lisossomos preenchidos com enzimas de degradação aparecem como pequenos vacúolos no citoplasma. Os monócitos se originam dos pró-monócitos, que são precursores de divisão rápida na medula óssea. Quando as células maduras entram no sangue periférico, elas são chamadas de monócitos; quando deixam o sangue e se infiltram nos tecidos elas sofrem mais alterações e são então conhecidas como macrófagos. Outras células derivadas desta linhagem incluem as células de Kupffer, macrófagos alveolares, microglia e osteoclastos.
Figura 3. Um monócito (célula grande à esquerda), que pode alcançar 17 µm de diâmetro, tem abundante citoplasma basofílico e um grande núcleo excêntrico.
Os macrófagos contêm receptores padrão de reconhecimento (receptores Toll-like [TLRs] 1 a 10)10 e receptores removedores. Além disso, os receptores para anticorpo e complemento potencializam sua capacidade de fagocitar organismos que são revestidos com estas substâncias. Os receptores de anticorpos reconhecem a porção Fc da IgG1, IgG3 e IgE. Existem dois receptores de complemento, CR1 e CR3. CR1 tem alta afinidade pelo componente C3b do complemento e tem uma menor afinidade pela iC3b e C4b. O CR3, também chamado de antígeno-1 do macrófago (MAC-1), interage com iC3b assim como com algumas moléculas de carboidrato, incluindo antígenos contendo carboidrato do protozoário Leishmania. Por meio destes receptores, os macrófagos atuam como células efetoras, atacando microrganismos e células neoplásicas e removendo material estranho.
Da mesma importância, os macrófagos apresentam o antígeno processado aos linfócitos e assim possuem um papel importante na indução das respostas imunológicas adquiridas. Uma pequena quantidade de antígeno da classe II do CMH está presente nos monócitos e sua expressão aumenta muito quando os macrófagos são ativados. Os macrófagos podem ser ativados por várias citoquinas, incluindo IFN-gama, fator estimulante de colônia de macrófagos e granulócitos (GM-CSF), fator ativador de macrófago (MAF) e fator inibidor de migração (MIF) [ver Mecanismos de resposta imunológica]. As citocinas como a IL-4 e o fator beta de crescimento transformador (TGF-beta) antagonizam esta ativação.
Os próprios macrófagos produzem várias substâncias solúveis que são importantes na resposta imunológica e no processo de inflamação. Estas substâncias incluem enzimas como o fator de ativação de plasminogênio e elastase; fatores de crescimento como GM-CSF; citocinas como IL-1, IL-6, IL-10, IL-12 e fator de necrose tumoral alfa (FNT-alfa); fatores que são críticos para combater microrganismos, como os metabólitos do oxigênio e óxido nítrico; componentes do complemento para as vias clássica e alternativa; MIPs e fatores que promovem o reparo tecidual, como o fator de crescimento de fibroblasto (FCF).
As células dendríticas são as CAPs profissionais que empregam respostas específicas por meio das células T.11-13 Elas estão presentes onde os antígenos e microrganismos têm o primeiro contato com o corpo, por exemplo, na pele (células de Langerhans) e os tratos gastrintestinal e respiratório. A função das células dendríticas é discutida adiante [ver Nódulos linfáticos e baço].
O sistema imunológico possui vários órgãos linfoides, incluindo timo, nódulos linfáticos, baço e amídalas, agregados de tecido linfoide nos órgãos não linfoides, como as placas de Peyer no intestino, grupos de células linfoides dispersas pelos tecidos conectivo e epitelial do corpo; assim como por toda a medula óssea e sangue e uma variedade de células individuais que viajam a partir de vários órgãos linfoides para o resto do corpo. Os linfócitos são derivados de precursores na medula óssea: as células T se desenvolvem no timo e são então exportadas para a periferia, enquanto as células B e NK se desenvolvem na medula óssea e então saem para a periferia. Das células hematopoiéticas não linfoides, os monócitos, macrófagos e células dendríticas são elementos importantes da imunidade inata e adquirida, enquanto os granulócitos (p. ex., neutrófilos, eosinófilos, basófilos e mastócitos) possuem papéis auxiliares no sistema imunológico.
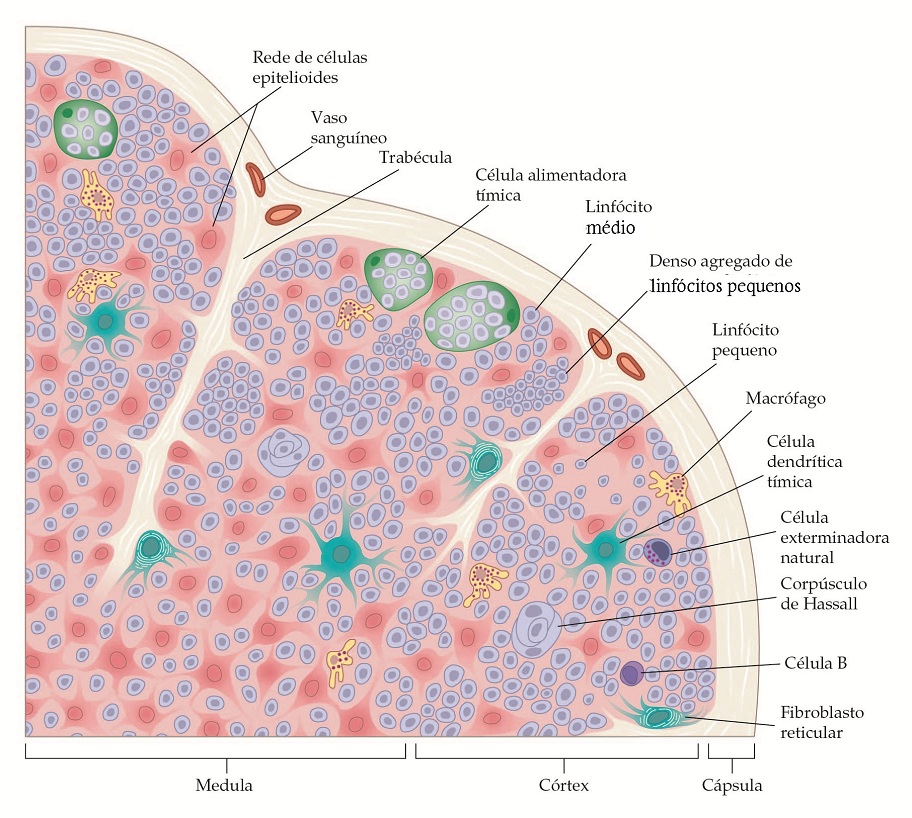
O timo, que se origina das terceira e quarta bolsas faríngeas do embrião, encontra-se no mediastino anterior e possui muitos lóbulos, cada um com um córtex e uma medula [Figura 4]. Os precursores da célula T derivados da medula óssea entram no timo, sendo os primeiros na área subcapsular. Eles se desenvolvem em células que expressam o complexo TCR-alfa-beta–CD3 e consequentemente adquirem o potencial para reagir com diferentes peptídeos ligados ao CMH. Esses timócitos C4+, CD8+ sofrem seleção negativa ou positiva, que envolve mecanismos complexos [ver Desenvolvimento do Repertório de Células T, adiante]. Apenas uma pequena percentagem de timócitos positivamente selecionados, CD4+ (que identificam classe II do CMH) ou CD8+ (que reconhecem a classe I do CMH), migra para a medula e então se desloca para o sistema linfoide periférico. Não está claro se as células T gama-delta se diferenciam no timo.
As células T maduras emigram pela parede das vênulas pós-capilares da medula para entrar na corrente sanguínea e subsequentemente residem nos órgãos periféricos do sistema linfoide. Uma vez nestes órgãos, os linfócitos deixam a corrente sanguínea; mais uma vez pelas vênulas pós-capilares, e viajam para as regiões das células T nos órgãos linfoides periféricos. Estes órgãos incluem o córtex interno dos nódulos linfáticos, bainhas periarteriais do baço e áreas intranodulares nas placas de Peyer, as amídalas e o apêndice. Acredita-se que algumas das células T presentes na mucosa intestinal (linfócitos intraepiteliais) se diferenciam fora do timo.
Pessoas que nascem sem o timo apresentam linfocitopenia com marcante depleção ou ausência de células T. As zonas das células T do sistema linfoide periférico também não apresentam linfócitos. A pessoa apresenta marcante comprometimento da imunidade mediada por células e as respostas de anticorpos que exigem a cooperação das células T (exceto a resposta da IgM) estão gravemente comprometidas [ver Deficiências nas imunoglobulinas e na imunidade mediada por células]. O timo involui com a idade, o que pode explicar o desenvolvimento de deficiências do sistema imunológico nas pessoas idosas.
Figura 4. A glândula timo é composta de vários lóbulos. A maioria dos linfócitos no córtex é imatura, são células de divisão rápida que podem ser facilmente destruídas pela cortisona. Durante a maturação, eles se deslocam para a medula, onde se tornam imunocompetentes e resistentes a cortisona. A partir deste momento, eles migram para os órgãos linfoides secundários, incluindo os nódulos linfáticos e o baço. A divisão e a maturação das células são influenciadas pelas células epiteliais; os densos agregados destas células formam estruturas conhecidas como corpúsculos de Hassall.
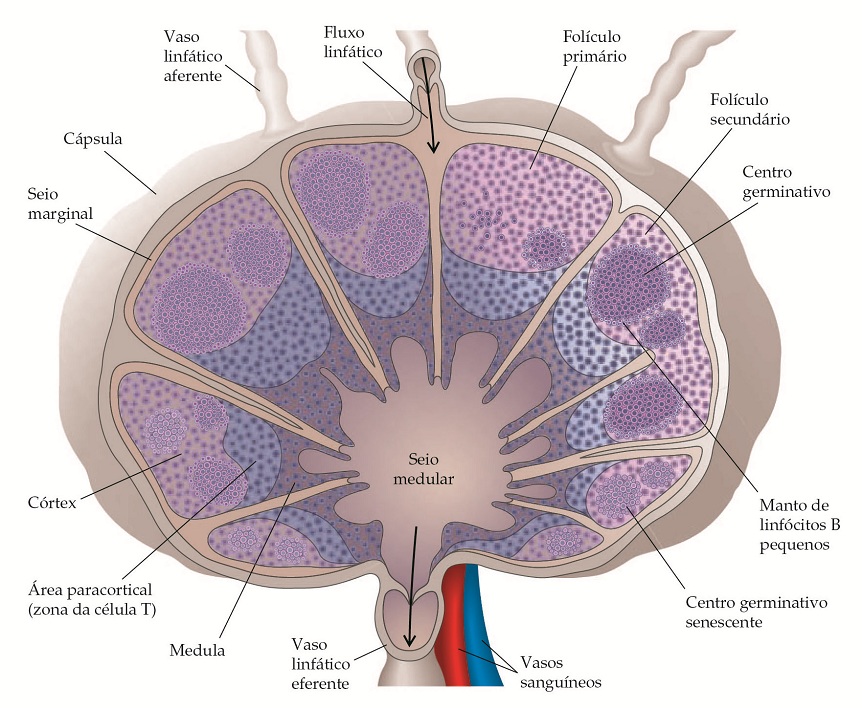
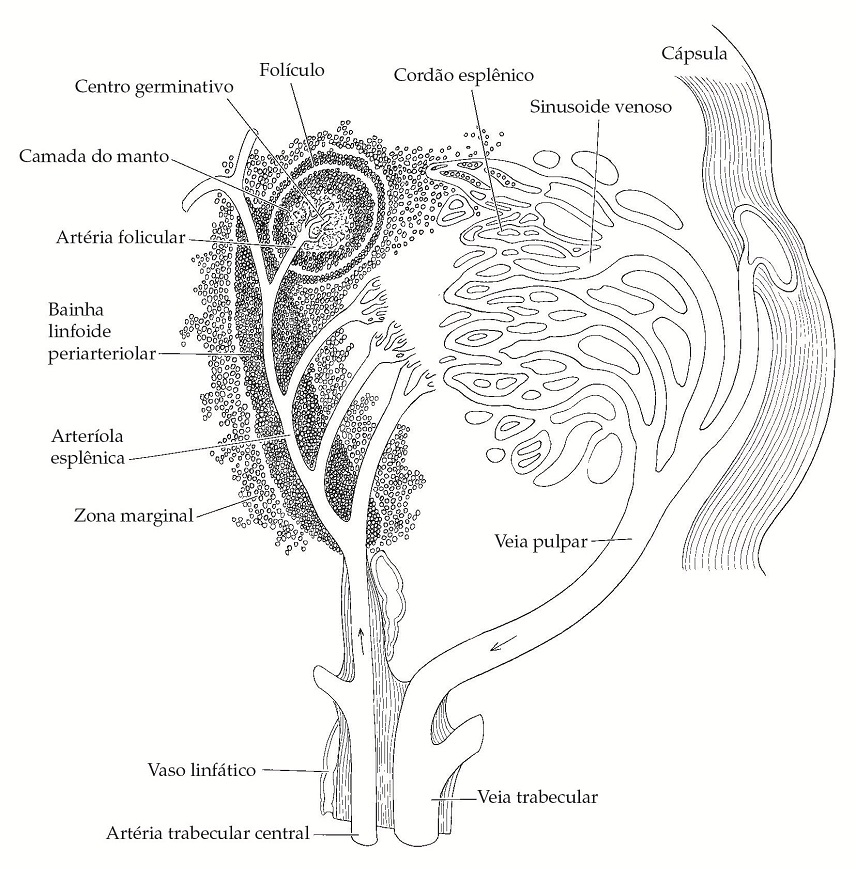
Os principais locais da ativação inicial das células T são os nódulos linfáticos e o baço, para onde convergem os linfócitos transportados no sangue, o antígeno transportado na linfa, alguns mediadores e células [Figura 5]. A linfa e as células entram no nódulo via região subcapsular, por meio de vasos linfáticos aferentes, infiltram no seio subcapsular e saem pelos vasos linfáticos eferentes no hilo. No baço, os linfócitos estão concentrados na polpa branca, que possui folículos com centros germinativos que circundam as arteríolas centrais [Figura 6].
Figura 5. Linfa contendo linfócitos, antígeno e mediadores solúveis drenados dos tecidos circundantes que entram nos nódulos linfáticos pelos vasos linfáticos aferentes. O nódulo linfático possui um córtex e uma medula. A área mais importante do córtex possui células B organizadas em folículos linfoides e sua área profunda, ou paracortical, consiste basicamente de células T e células dendríticas. Após as células B encontrarem os antígenos para os quais elas possuem receptores, junto com as células T adequadas, as áreas centrais da marcante proliferação de células B chamadas de centros de germinação se desenvolvem nos folículos linfoides. À medida que estas reações terminam, os centros germinativos se tornam senescentes. Os cordões medulares estão na medula, que são séries de macrófagos e células plasmáticas.
Figura 6. No baço, as células linfoides ao redor da arteríola esplênica formam a bainha linfoide periarteriolar, que é basicamente uma área dependente do timo. O folículo com um centro germinativo contém células B. As células T são encontradas principalmente na região central das bainhas linfoides periarteriolares, enquanto as células B nos centros germinativos se concentram mais no sentido da periferia das bainhas. Os linfócitos entram e deixam as bainhas linfoides periarteriolares por meio dos capilares das arteríolas centrais na zona marginal.
A infraestrutura do nódulo linfático é uma extensa rede reticular onde CAPs e células T se encontram e interagem. Por exemplo, as células dendríticas derivadas da medula óssea presentes na pele coletam o antígeno e viajam pelos vasos linfáticos para o nódulo linfático de drenagem. As células então migram, por meio do assoalho do seio subcapsular do nódulo linfático para as regiões interfoliculares, onde elas se acumulam na rede reticular do paracórtex como células dendríticas interdigitantes (IDCs). As células T do sangue migram pelas vênulas pós-capilares especializadas, conhecidas como vênulas endoteliais altas (HEV) e migram ao longo da mesma rede reticular, onde entram em contato com as numerosas IDC apresentadoras de antígenos.14
Os centros germinativos contêm células B, que são derivadas das células-tronco e se diferenciam na medula óssea [Figura 1]. Os folículos linfoides contêm células dendríticas foliculares (FDCs), que não são derivadas da medula óssea. As FDCs, encontradas apenas nos folículos linfoides, expressam os receptores do complemento CR1, CR2, CR3 e receptores Fc. Estes receptores permitem que as FDCs apresentem o antígeno às células B ativadas na forma de complexos antígeno-anticorpo-complemento. As FDCs podem reter estes complexos por muito tempo.
Os centros germinativos ativos são circundados por um manto de células B (células do manto folicular) que expressam IgD na sua superfície e podem amadurecer em células plasmáticas que produzem apenas anticorpos IgM. As células B no centro germinativo (centrócitos) sofrem uma troca de classe para produzirem outros isotipos como IgG, IgA e IgE. Acredita-se que as células B com anticorpos de alta afinidade nas suas superfícies são selecionadas ao se ligarem a FDCs retentoras de antígenos. As células B não selecionadas morrem de apoptose (morte celular programada). A célula B mais madura, a célula de memória, também é encontrada no centro germinativo e pode se desenvolver em células plasmáticas produtoras de todos os isotipos de anticorpos.
As células dendríticas derivadas da medula óssea, encontradas nas áreas paracorticais do nódulo linfático, são CAPs profissionais que têm papel crucial no início das respostas imunológicas dependentes das células T.12-14 Estas células também são encontradas como células imaturas nos órgãos não linfoides, especialmente na epiderme, onde são chamadas de células de Langerhans. Por meio de muitos de seus receptores, em particular TLR1, TLR10 e receptores removedores, as células dendríticas imaturas internalizam de forma eficiente, processam os antígenos e os apresentam via seu CMH. Existem dois subconjuntos de células dendríticas, mieloides e plasmacitoides, que produzem citocinas diferentes. Algumas citocinas (p. ex., IL-1-beta e TNF-alfa) promovem a migração das células dendríticas através dos vasos linfáticos aferentes para os nódulos linfáticos. Neste local, eles amadurecem, perdem sua capacidade de fagocitose e expressam moléculas coestimuladoras críticas CD80 (B7-1) e CD86 (B7-2); estas moléculas coestimuladoras potencializam sua capacidade de apresentar os antígenos às células T. As células dendríticas imaturas também são encontradas nos nódulos linfáticos, onde podem fagocitar o antígeno que entra via vasos linfáticos aferentes e então amadurecem em CAPs.
Os linfócitos também são encontrados em vários outros locais. O tecido linfoide associado ao intestino inclui as placas de Peyer e o apêndice. Estes tecidos linfoides contêm regiões com concentrações de células T ou B semelhantes às encontradas nos centros germinativos. Acredita-se que as células epiteliais especializadas, chamadas de células M, possuem uma capacidade única de captar e apresentar o antígeno para os linfócitos adjacentes.15 As células M também são encontradas próximas das placas de Peyer. Outros linfócitos presentes no intestino são os linfócitos da lâmina própria (LPLs), encontrados nos vilos e os linfócitos intraepiteliais (IELs), encontrados entre as células epiteliais. A migração e a aderência dos LPLs são, em parte, ditadas pelas integrinas e selectinas, que são moléculas de superfície nos tecidos e células que medeiam as interações celulares e células residentes. As células linfoides associadas com a mucosa também são encontradas no trato respiratório e no trato geniturinário. O sistema imunológico especializado da pele contém células de Langerhans na epiderme (células dendríticas mieloides especializadas) e uma maior concentração de células T gama-delta do que em outro local.16
Existem 3 principais tipos de circulação de linfócitos:
a semeadura das células-tronco a partir do fígado ou medula óssea fetal para os principais órgãos linfoides e a subsequente diferenciação e distribuição destas células para o sistema linfoide periférico;
a recirculação de linfócitos do sangue para linfa e vice-versa;
a distribuição das células efetoras para partes específicas do corpo.
Os linfócitos circulam continuamente do sangue para os tecidos e de volta para o sangue. Entretanto, o trânsito das células T jovens (CD45RA) é diferente do das células efetoras ou de memória ativadas (CD45RO). As células T jovens recirculam através dos tecidos linfoides secundários, como os nódulos linfáticos, baço, amídalas e placas de Peyer para microambientes especiais onde elas encontram antígeno, citocinas e outras células que levam a sua ativação. Por outro lado, o efetor ativado ou as células de memória também podem transitar para os locais extralinfoides em vários tecidos, como a pele ou a lâmina própria do intestino.17
O deslocamento dos linfócitos para o endotélio vascular e sua passagem por este são controlados pela expressão de vários receptores na superfície dos linfócitos e contrarreceptores no endotélio vascular. Para interromper o fluxo de células no sangue e nos vasos linfáticos, ocorre a adesão inicial primária entre os receptores dos linfócitos nas microvilosidades das células, como L-selectina e o contrarreceptor no endotélio, como a adressina do nódulo linfático periférico (PNAd). Outros receptores celulares permitem a fixação a E-selectina e P-selectina endoteliais. Subsequentemente, as células podem se fixar e girar usando os receptores de superfície integrina-Ig como alfa-4-beta-7 e alfa-4-beta-1, que se ligam à molécula de adesão celular adressina da mucosa endotelial (MAdCAM-1) e à molécula de adesão celular vascular (VCAM-1), respectivamente. Estas interações podem levar a uma parada estável envolvendo um receptor que aciona a adesão via sinalização intracelular de uma proteína de ligação de trifosfato de guanosina (GTP). A cooperação entre as interações do receptor é essencial porque a interação inicial com L-selectina pode ser fraca demais para induzir a interação estável LFA-1/ICAM-1 (antígeno-1 associado com a função do leucócito/molécula-1 de adesão intracelular) e, portanto, requer a interação alfa-4-beta-7/MAdCAM-1. Por outro lado, quando os tecidos apresentam altos níveis do receptor L-selectina, o contato e a rotação medidos pela L-selectina podem ser suficientes para permitir uma parada estável mediada pelo LFA-1. Os linfócitos podem, então, atravessar o endotélio vascular, um processo chamado de diapedese. Um processo semelhante está envolvido com a diapedese de outros leucóticos.17
Acredita-se que as variações nestes mecanismos canalizam os subconjuntos de linfócitos para os diferentes microambientes nos nódulos linfáticos, como os centros germinativos, as áreas paracorticais e a zona T, onde células, antígeno e fatores solúveis levem a respostas imunológicas particulares. Estes microambientes são também regulados por várias citocinas, como TGF-beta-1, que pode sobrerregular as integrinas e mediar a ligação das células B às CAPs. As citocinas como TNF-alfa regulam os receptores de adesão dos linfócitos e outras citocinas influenciam a atividade dos linfócitos à medida que eles atravessam os tecidos. Algumas quimiocinas, como RANTES, também ativam a expressão de moléculas de adesão na superfície das células T efetoras. Outras quimiocinas, como a quimiocina expressa pelo timo (TECK), recrutam células T que carregam receptores TECK (CCR9) para seu órgão residente, neste caso, o intestino.18,19
As respostas imunológicas são controladas por 3 grandes famílias de genes:
os genes que codificam elementos variáveis das imunoglobulinas;
os genes que codificam os receptores de células T (T Cell Receptors - TCRs);
os genes que codificam os antígenos do CMH [ver Antígenos, anticorpos e receptores das células T].
Cada pessoa possui um número incrível de genes que codificam elementos diversos das imunoglobulinas e TCRs, o que permite o reconhecimento específico de milhões de antígenos. Entretanto, a grande variabilidade do CMH aplica-se a população como um todo, qualquer pessoa terá apenas algumas poucas variações.
A capacidade das células T e anticorpos produzidos pelas células B em distinguir entre os antígenos é regulada por dois conjuntos independentes de genes de regiões variáveis para as células T e B, cada um composto por segmentos de DNA reorganizados em V (variável), D (diversidade) e J (união). A reorganização das sequências de DNA destes genes ocorre à medida que as células T e B amadurecem, dando origem a TCRs e imunoglobulinas. Ocorre mutação somática adicional nas células B durante sua maturação, o que expande o repertório. Este sistema pode distinguir bilhões de antígenos [ver Antígenos, anticorpos e receptores das células T].
Cada linfócito tem um receptor de superfície que reconhece um único determinante antigênico ou epítopo. Os receptores da célula B reconhecem antígenos nativos. Após as células B interagirem com um antígeno, elas se proliferam e se diferenciam em células plasmáticas para produção de anticorpos e algumas se tornam células de memória.
Antes de ser reconhecido pelas células T, um antígeno é captado por uma CAP (p. ex., uma célula dendrítica ou um macrófago) que quebra o antígeno em pequenos fragmentos de peptídeos. Na CAP, alguns fragmentos de peptídeo ou epítopos são recolhidos por moléculas da classe II do CMH e transportados para a superfície da CAP. Assim, os TCRs não reconhecem o antígeno nativo, apenas suas partes processadas [ver Antígenos, anticorpos e receptores das células T].
A exigência de que o antígeno seja apresentado junto com uma molécula da classe I ou II do CMH é conhecida como restrição do CMH. A natureza do CMH explica por que algumas pessoas podem não responder a alguns antígenos. Por exemplo, embora os TCRs reconheçam epítopos ligados a moléculas do CMH, alguns peptídeos do antígeno podem não se encaixar na fenda da molécula do CMH particular de uma pessoa. Desta forma, o tipo adequado de célula T não reagirá com esse epítopo e a pessoa não será capaz de desenvolver uma resposta imunológica contra ele.
Dos dois tipos de células T, alfa-beta e gama-delta, apenas o desenvolvimento das células que expressam TCR-alfa-beta está bem elucidado. Após a reorganização dos genes, o TCR-alfa-beta é expresso na superfície do timócito cortical imaturo junto com as proteínas CD3 [Figura 1]. Ao mesmo tempo em que ocorre a reorganização dos genes, CD4, CD8 e CD3 são expressas.6 Timócitos maduros positivos únicos (CD4+ ou CD8+) carregando altos níveis de TCR-alfa-beta são selecionados do conjunto de timócitos CD4+, CD8+ por meio de processos chamados de seleção positiva e seleção negativa.
A seleção positiva é controlada pelas células epiteliais do córtex do timo e CAPs dedicadas, como macrófagos, células dendríticas e celular interdigitantes. Muitas destas células do estroma estão localizadas na junção corticomedular. Como as células T podem reagir a antígenos apenas na associação com auto-CMH, apenas células T com um TCR que pode se ligar a auto-CMH são selecionadas. Quando estas células reagem com auto-CMH nas células do estroma do timo, células CD4+, CD8+ TCR-alfa-beta que se ligam a moléculas da classe II do CMH se tornam CD4+ e CD8–, subregulando CD8 e suprarregulando o complexo TCR-CD3. Por outro lado, as células CD4+ CD8+ que se ligam a moléculas da classe I do CMH subregulam CD3 e tornam-se células CD4–, CD8+ TCR-alfa-beta+. Desta forma, são selecionadas as células auto-CMH-restritas CD4+ TCR-alfa-beta e células CD8+ TCR-alfa-beta. Apenas timócitos cujo TCR tenha uma afinidade moderada pelo peptídeo do CMH são selecionados de forma positiva. As células CD4 ou CD8 contribuem para a avidez da interação já que elas próprias ligam-se à classe II e à classe I do CMH, respectivamente.
A maioria das células T não interage com auto-CMH e, portanto, sofre apoptose por causa da ausência de interação. O processo ativo da seleção negativa elimina células T que possuem um TCR com uma alta avidez pelo peptídeo do CMH. Se não forem removidas, estas células T podem provocar grave doença autoimune. Existem muitos autoantígenos nas células epiteliais do timo e os estudos mostram que alguns autoantígenos podem ser apresentados por várias CAPs no timo, como macrófagos e células dendríticas. É possível que a seleção negativa para alguns autoantígenos ocorra quando as células T se deslocam para o sistema linfoide periférico após deixarem o timo. Esta interação de alta afinidade entre o autoantígeno apresentado pelo CMH e o TCR nas células T imaturas dispara vários processos que levam à morte celular (p. ex., apoptose) (ver adiante).
A seleção positiva das células B precursoras ocorre na medula óssea. As células pré-B possuem na sua superfície a cadeia pesada de IgM (µ) em associação com a cadeia leve substituta (lambda 5 ou ?5). Se o último receptor não estiver na superfície, a célula é eliminada por apoptose. As células pré-B se desenvolvem em células B imaturas, que carregam IgM na sua superfície. Estas células se desenvolvem em células B maduras exceto se forem eliminadas pelo reconhecimento do antígeno e seleção negativa nos nódulos linfáticos. Se a afinidade for alta, a eliminação envolve apoptose; se a afinidade for baixa, ela envolve anergia (a célula está presente, mas não se desenvolve).
A remoção de células T e B autorreativas na periferia também usa apoptose. Os sinais da apoptose que atuam na membrana celular levam à morte celular programada com condensação e ruptura da cromatina e, dentro de minutos a horas, a célula é destruída e depurada pelos macrófagos. Os principais receptores nas células linfoides que disparam a apoptose são 18 membros da família de receptores TNF.20 Os exemplos incluem o receptor Fas (CD95/APO-1), que é acionado pelo ligante Fas e o receptor-1 de TNF (TNFR-1), acionado por TNF-alfa e linfotoxina-alfa. Os TNFRs também acionam vias que podem ativar o fator nuclear ?B (TNF-?B) que protege contra a apoptose.21 Os sinais para apoptose eventualmente atuam numa família de proteases derivadas da cisteína semelhantes a enzima conversora de IL-1b (ICE), o protótipo que atua no IL-1b precursor da citocina, convertendo-a na citocina ativa. As proteases relacionadas a ICE ativam outras proteases, que então atuam em diversos substratos, o que, por sua vez, leva a apoptose.
A característica da resposta imunológica é sua capacidade de aumentar o número de linfócitos específicos para o antígeno após ocorrer um estímulo antigênico; por causa das células T e B de memória, a exposição posterior ao mesmo antígeno leva a uma resposta mais rápida e maior (ou seja, resposta anamnéstica). A base para esta resposta potencializada é a proliferação de linfócitos específicos para antígeno e a produção de células de memória após os linfócitos interagirem com um antígeno. Estas respostas são mediadas pela produção de citocinas pelos linfócitos e outras células [ver Mecanismos da resposta imunológica]. Os mecanismos de resposta imunológica também são amplificados pela liberação de substâncias mediadoras a partir de mastócitos e basófilos revestidos por anticorpos, da ativação de proteínas do complemento e da expressão das moléculas de integrina nas células. A permeabilidade vascular alterada, a expressão dos receptores pelos leucócitos nas células endoteliais e a liberação de fatores quimiotáticos por meio destes mecanismos secundários atraem muitos outros tipos de células para a reação. Estas células contribuem muito para a inflamação resultante ao auxiliar o processo de fagocitose e descarte de antígenos estranhos.
Após as células T interagirem com um antígeno, elas se proliferam e se diferenciam em células efetoras, algumas células T auxiliares e outras células citotóxicas (CTLs); todas possuem um componente da célula de memória.
Uma linhagem independente e células CD4+ são células T reguladoras (Treg), que controlam as respostas imunológicas ao produzirem citocinas inibidoras e ao direcionarem o contato célula a célula.22,23 As células Treg específicas a antígenos também se desenvolvem como uma linhagem independente no timo.24 Os aspectos da memória das células Treg são desconhecidos.
As células B da memória potencializam a resposta imunológica ao antígeno anteriormente encontrado como parte da resposta anamnéstica. Estas células B de memória sofrem mutação somática nas regiões variáveis dos seus genes para imunoglobulina. Quando esta mutação somática ocorre nos centros germinativos dos nódulos linfáticos que contêm antígenos ligados a células dendríticas foliculares, ela leva à seleção de células B de memória que possuem receptores de alta afinidade para os antígenos.
Os autores não possuem relações comerciais com os fabricantes dos produtos ou fornecedores de serviços discutidos nesta subseção.
1. Burnet M: The Clonal Selection Theory of Acquired Immunity. Cambridge University Press, London, 1959
2. Mosmann TR, Coffman RL: Heterogeneity of cytokine secretion patterns and functions of helper T cells. Adv Immunol 46:111, 1989
3. Feng Y, Broder CC, Kennedy PE, et al: HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein–coupled receptor. Science 272:872, 1996
4. Choe H, Farzan M, Sun Y, et al: The ß-chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell 85:1135, 1996
5. Doranz BJ, Rucker J, Yi Y, et al: A dual-tropic primary HIV-1 isolate that uses fusin and the ß-chemokine receptors CKR-5, CKR-3, and CKR-2 as fusion cofactors. Cell 85:1149, 1996
6. CD Index. PROW (Protein Reviews on the Web). National Center for Biotechnology Information, 2002 www.ncbi.nlm.nih.gov/prow/guide/45277084.htm
7. Qin XF, Schwers S, Yu W, et al: Secondary V(D)J recombination in B-1 cells. Nature 397:355, 1999
8. Natarajan K, Dimasi N, Wang J, et al: Structure and function of natural killer cell receptors: multiple molecular solutions to self, nonself discrimination. Annu Rev Immunol 20:853, 2002
9. Vilches C, Parham P: KIR: diverse, rapidly evolving receptors of innate and adaptive immunity. Annu Rev Immunol 20:217, 2002
10. Janeway CA Jr, Medzhitov R: Innate immune recognition. Annu Rev Immunol 20:197, 2002
11. Mellman I, Steinman RM: Dendritic cells: specialized and regulated antigen processing machines. Cell 106:255, 2001
12. Rissoan MC, Soumelis V, Kadowaki N, et al: Reciprocal control of T helper cell and dendritic cell differentiation. Science 283:1183, 1999
13. Siegal FP, Kadowaki N, Shodell M, et al: The nature of the principal type 1 interferon-producing cells in human blood. Science 284:1835, 1999
14. Liu YJ, Kanzler H, Soumelis V, et al: Dendritic cell lineage, plasticity and cross-regulation. Nat Immunol 2:585, 2001
15. Neutra MR, Mantis NJ, Kraehenbuhl JP: Collaboration of epithelial cells with organized mucosal lymphoid tissues. Nat Immunol 2:1004, 2001
16. Randolph GJ: Is maturation required for Langerhans cell migration? J Exp Med 196:413, 2002
17. Steeber DA, Tedder TF: Adhesion molecule cascades direct lymphocyte recirculation and leukocyte migration during inflammation. Immunol Res 22:299, 2000
18. Rossi D, Zlotnik A: The biology of chemokines and their receptors. Annu Rev Immunol 18:217, 2002
19. Sallusto F, Mackay CR, Lanzavecchia A: The role of chemokine receptors in primary, effector, and memory immune responses. Annu Rev Immunol 18:593, 2000
20. Gupta S: A decision between life and death during TNF-alpha–induced signaling. J Clin Immunol 22:185, 2002
21. Beg AA, Baltimore D: An essential role for NF-.B in preventing TNF-a–induced cell death. Science 274:782, 1996
22. Shimizu J, Yamazaki S, Takahashi T, et al: Stimulation of CD25(+)CD4(+) regulatory T cells through GITR breaks immunological self-tolerance. Nat Immunol 3:135, 2002
23. Sakaguchi S: Regulatory T cells: key controllers of immunologic self-tolerance. Cell 101:455, 2000
24. Jordan MS, Boesteanu A, Reed AJ, et al: Thymic selection of CD4+CD25+ regulatory T cells induced by an agonist self-peptide. Nat Immunol 2:301, 2001
Figuras 1, 4 e 5: Seward Hung.
Figura 6: Carol Donner.
O MedicinaNET é o maior portal médico em português. Reúne recursos indispensáveis e conteúdos de ponta contextualizados à realidade brasileira, sendo a melhor ferramenta de consulta para tomada de decisões rápidas e eficazes.
| Medicinanet Informações de Medicina S/A Cnpj: 11.012.848/0001-57 | info@medicinanet.com.br |
MedicinaNET - Todos os direitos reservados.
Em função da pandemia do Coronavírus informamos que não estaremos prestando atendimento telefônico temporariamente. Permanecemos com suporte aos nossos inscritos através do e-mail info@medicinanet.com.br.